激活小麦基因组中的抗病基因(前沿研究优化关键物候基因)
本文选自中国工程院院刊《Engineering》2018年第4期
作者:M.A.N. Nazim Ud Dowla,Ian Edwards,Graham O’Hara,Shahidul Islam,Wujun Ma
来源:M.A.N. Nazim Ud Dowla,Ian Edwards,Graham O’Hara,Shahidul Islam,Wujun Ma.Developing Wheat for Improved Yield and Adaptation Under a Changing Climate: Optimization of a Few Key Genes[J].Engineering,2018,4(4):514-522.
导 语
谷类作物构成了全球人类主要的食物来源,其中,小麦是继水稻之后世界第二大最重要的粮食作物,是世界上种植面积最大的谷类作物。受气候变化带来的气温和降水影响,干旱对小麦产量的影响将会更加恶化。
中国工程院院刊《Engineering》刊发的《气候变化条件下提高小麦产量和适应性——关键物候基因的优化 》中指出,由于物候基因的多样性,小麦具有广泛的适应性。文章综述了春化(Vrn)、光周期(Ppd)和矮秆(Rht)基因等小麦物候学研究的最新进展。在不同气候条件下Vrn和Ppd基因的等位基因、单倍型和拷贝数有不同的反应,因而不仅可以改变发育阶段,而且可以提高产量。与模式植物拟南芥相比,还有更多尚未发现的小麦物候基因,在目标环境中量化其效应将有助于通过育种提高小麦的耐旱性。因此,通过配置适当的物候基因、Rht基因和其他耐旱性相关的重要生理性状的组合,可以在水资源有限的环境中提高小麦产量。

一、引言
谷类作物构成了全球人类主要的食物来源。其中,小麦(Triticum aestivum L.)是继水稻之后世界第二大最重要的粮食作物,是世界上种植面积最大的谷类作物。小麦是粮食安全的中心支柱之一,为全世界人口提供20%的总热量和相近比例的总蛋白质。全球小麦平均产量为3.3 t·hm–2;但不同区域平均值差异很大,据2015年联合国粮农组织数据库(The Food and Agricuture Organization Corporate Statistical Database,FAOSTAT)显示,澳大利亚为1.7 t·hm–2,而世界上一些高产地区能达到9 t·hm–2。
产量损失通常是由于不同的环境情况造成的,减产幅度可达69.1%。在大多数发达国家,小麦主要种植在雨养边缘地区,降雨量不足和不稳定极大地限制了小麦产量(表1)。干旱是全球在大约6.5×10 7t·hm–2的耕地上制约小麦产量的关键性胁迫因素,减产幅度高达50%。模拟试验表明,在灌溉条件下,边缘小麦生长环境中的水分胁迫可使小麦产量潜力降低50%~90%。2012年,全球小麦总产量下降1.4%,主要原因是美国、欧洲和中亚地区发生了严重干旱(数据来自2012年FAOSTAT数据库)。2006年,澳大利亚小麦产量比过去50年下降了46%,造成小麦产业损失数十亿美元(数据来自2013年FAOSTAT数据库)。
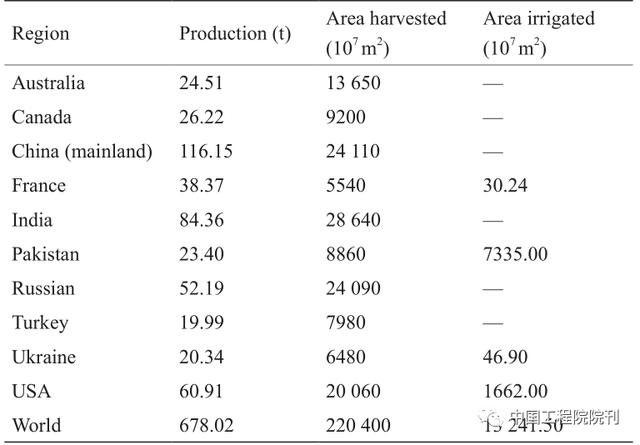
表1 世界主要小麦生产国:5年(2008—2012年)的小麦产量、收获 面积和灌溉面积的平均值
由于气候变化对气温和降水的影响,预计未来干旱对小麦产量的影响将会更加严重。据估计,在过去的29年里,气温上升1℃将导致小麦比没有全球变暖影响时减产6%。根据政府间气候变化专门委员会(The Intergovernmental Panel on Climate Change,IPCC)的第5次评估报告,到21世纪末,全球平均气温将上升3.7℃,酷暑和寒夜的天数将比现在多50%。降水年型的变化加上温度升高将影响世界主要作物的生产,除非采取限制全球变暖的保护措施、种植合适的品种或采取适当的作物管理措施,否则到2050年小麦产量可能将下降23.2%~27.2%。
与高纬度地区相比,低纬度地区的小麦产量会更容易受3~5℃升温的影响,而温度升高2℃,产量可能会下降40%。澳大利亚小麦带属于典型的地中海气候,降水多发生在冬季,春季降雨较少,夏季是最干旱的季节。因此,春季水分胁迫是限制这些地区产量提高的主要因素,并且常常与拔节期、开花期和籽粒灌浆期相一致。在这些环境中,后期籽粒灌浆期高温常常与干旱相伴而生,进一步限制籽粒产量。IPCC在其第5次评估报告中预测,到2030年、2050年和2070年,年平均气温将上升2.2~5℃,降水年型发生 5%~–30%的变化,将导致受旱地区分别扩大5.4%、4.6%和3.8%。在这种情况下,植物育种者必须做好迎接气候变化挑战的准备,通过培育更加适应水资源有限环境的品种来养活世界。更好地利用小麦现有的遗传资源对于保持水资源有限的环境中的小麦产量潜力并使其最大化至关重要,物候优化是实现这一目标的最有效途径之一。
物候是作物适应特定环境的关键因素,正确理解物候特性的遗传控制将使育种者培育出更能适应特定环境的品种。有证据表明,干旱造成的产量损失取决于其发生的生长期,以及胁迫的持续时间和强度。开花期的穗重与籽粒产量正相关,并且可以在不影响其他发育阶段的情况下加以操作,穗发育(从小穗分化至开花期)是决定籽粒产量最重要的阶段。因此,通过确保在最敏感的发育阶段不发生胁迫,将干旱的不利影响降到最低。因此,开花的微调和发育阶段的持续时间被认为可使小麦更好地适应水资源有限的环境,或者回避这些限制因素。物候基因也调控小麦的生理发育,一些形态生理性状已被认为可有效地培育适应干旱条件的品种。鉴于很多重要性状及其在胁迫环境中的互作,熟悉水分胁迫的遗传控制和生理基础将有助于提高水分受限环境下的产量。
笔者也认识到良好的农事作业(管理)和其他几个涉及生理机制的性状在减少干旱不利影响方面的重要性,但本文的重点在于阐述作为避免干旱胁迫最重要的措施之一的物候基因(图1)。因此,本文力图总结迄今在关键物候基因研究方面所取得的进展,并将这些知识整合到适应未来气候变化的新品种培育工作中。此外还尝试汇集鉴定Vrn(春化)和Ppd(光周期)基因的等位基因的分子标记信息,这些信息有助于在目标环境下的品种的评价以及标记辅助育种。
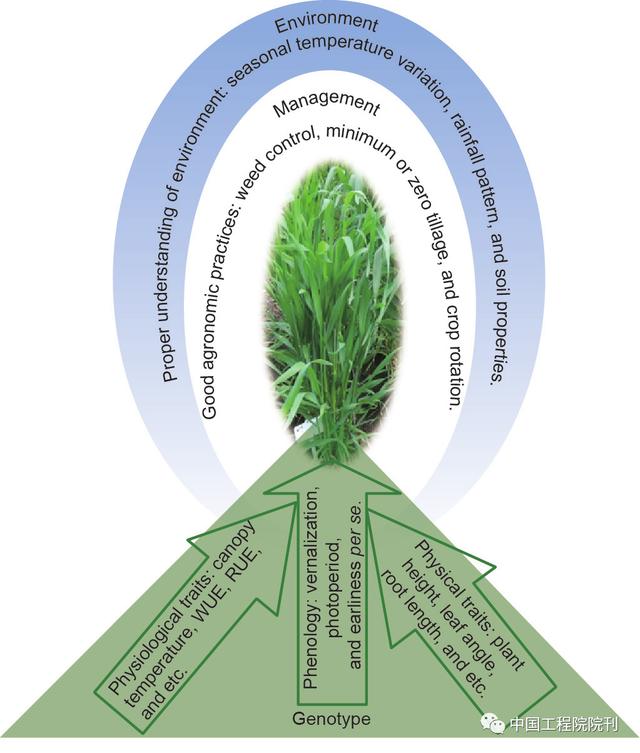
图1 抗旱盾牌:在水资源有限的环境中维持小麦产量的补充方法。WUE:水利用效率;RUE:辐射利用效率
二、物候基因
小麦适应广泛的农业环境。开花与较为广泛气候条件的同步性在很大程度上受控于:Vrn基因(暴露于低温的要求)、Ppd基因(光周期敏感性)和早熟性本身(Eps)基因。因此,基因型对特定环境的适应取决于这三组基因的相互作用。
(一)春化基因
春化通过在茎尖分生组织中诱导来自叶原基的花原基促进植物从营养生长期向生殖生长期转换。小麦有3个基因决定春化要求:Vrn1、Vrn2和Vrn3。
3个直系同源的Vrn1基因Vrn-A1、Vrn-B1和Vrn-D1分别位于普通小麦的部分同源染色体5A、5B和5D的长臂,主要控制春化要求。Vrn2也位于5A染色体长臂,Vrn3位于7B染色体短臂。冬小麦品种需要一定时间的低温来诱导开花,而无需春化即可开花的品种则被称为春性品种。Vrn-A1、Vrn-B1、Vrn-D1和Vrn3的显性等位基因控制春性生长习性,因此,3个Vrn1基因座任何一个显性等位基因都表现春性品种。
另一方面,Vrn2控制冬性品种,并且对于Vrn1显性等位基因表现上位性。Vrn2是一种延迟开花的花阻遏蛋白,但长日照条件下的春化抑制了Vrn2的表达而增强了Vrn1的表达。已经发现了Vrn1的多个对春化作用产生不同水平响应的等位基因,并且明确了其对开花的影响(表2),这些等位基因具有适应值。开花的程度取决于Vrn1表达的基础水平,并且Vrn1的一些等位基因在没有预先低温处理的情况下可以表达,因此花可以在未经春化的情况下开放。Vrn1基因第一个内含子中的启动子突变或者缺失可导致Vrn1未经春化而表达,Vrn1基因的启动子突变或其第一个内含子的缺失可导致Vrn基因未经春化而表达,缺失越大的等位基因在未经春化的开花早期越活跃。另一方面,当缺少Vrn2基因的功能拷贝时小麦和大麦品种不经春化就可早开花。在Vrn2缺失条件下,Vrn3基因也可高水平表达,无论日长或春化与否,Vrn3的活性等位基因都可加速开花。因此,3个Vrn基因可以通过控制世界各地小麦品种春化需求来影响开花。

表2 Vrn1和Ppd1基因座已知的等位基因现状
(二)光周期基因
小麦是一种长日照植物,需要长日照条件(>14 h光照)才能开花,而光周期不敏感的品种在短日照条件下(10 h或更少光照)也能开花。这种光周期敏感性由第2部分同源群染色体短臂上的半显性部分同源基因Ppd1控制,与Vrn1一样,显性等位基因光周期不敏感。Worland在不同小麦生产区域对光周期不敏感的等位基因Ppd1的作用进行了14年的全面研究,发现不敏感的Ppd1使开花期提前了9~15 d,这种早熟性可以在水资源有限的环境中通过避免干旱提高产量。早熟的Ppd1基因也有一些多效的作用,包括降低株高及减少分蘖数和每穗小穗数。然而,提高小穗的育性可以补偿产量损失。显然,Ppd1不敏感性使小穗形成终止的时间提前,从而通过减少小穗数而使开花期提前。但是,这并不影响叶原基和花原基产生率。3个Ppd1a基因座的效能也有差异,具有Ppd-A1a和Ppd-D1a的植株比具有Ppd-B1a的植株开花更早。与已经鉴定出许多Vrn-1等位基因的方式相同,最近还在普通小麦和硬粒小麦中发现Ppd-1基因的所有3个同源基因座的许多等位基因及其单倍型(表2),在育种计划中具有重要的利用价值。
(三)早熟性本身基因(Eps)
Eps基因控制与温度和光周期无关的开花期。迄今为止,在小麦中很少发现Eps基因,但是一些数量性状基因座(QTL)研究表明,大多数部分同源群染色体携带了这些基因,它们作为QTL效应而非Ppd和Vrn途径中的主要基因。Eps基因参与开花期的微调,因此可用于适应特定的气候条件。
三、物候基因的分子干预
与拟南芥的研究一致,在分子水平上阐明小麦开花途径方面已经取得重要的进展。分子和序列分析显示Vrn1编码类似于拟南芥分生组织特征基因APETA-LA1(AP1)、CAULIFLOWER(CAL)和FRUITFULL(FRU)的MADS-box转录因子,调节茎尖分生组织,确定从营养生长到生殖生长的转变。启动子区域的插入、缺失和突变与Vrn1的等位基因变异有关。在此之后,一系列分子标记被开发出来(表3),并成功地用于鉴定当地小麦品种以及CIMMYT(International Maize and Wheat Improvement Center)种质中这些等位基因的频率。Vrn2编码锌指CCT域转录因子,是一种花的阻遏物,受春化处理和短日照的限制。Vrn2在拟南芥中的作用与FLOWERING LOCUS C(FLC)非常相似,但是实际上不是直系同源基因,说明春化途径是独立进化的。春化基因Vrn3与拟南芥FLOWERING LOCUS(FT)类似,其显性等位基因与TaFT启动子中一个逆转录因子的插入有关,导致早开花。最近通过对一些中国小麦品种的筛选又发现了Vrn3的两个显性等位基因,观察到因其作用而导致的80 d抽穗的变异。这些Vrn1基因的等位基因变异量化了春化作用,并通过与光周期基因Ppd1相互作用来确定开花时间。后者是伪反应调节因子(pseudo response regulator,PRR)基因家族的成员,其中,不敏感性与启动子区域内的缺失或转座子插入以及拷贝数变异相关。在小麦中,Ppd-1直接调控FlOWERING LOCUS T1(FT1),启动子缺失的突变导致FT1的过表达,进而导致早开花。已经开发了可以鉴别具有不同启动子缺失的Ppd-1突变体的分子标记(表2),这些突变体可以促进其对小麦开花时间的影响。
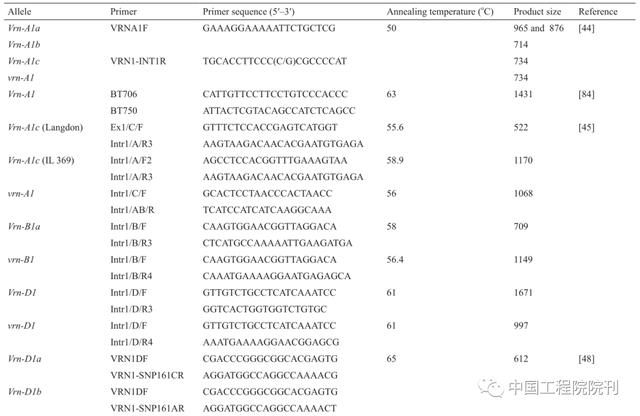
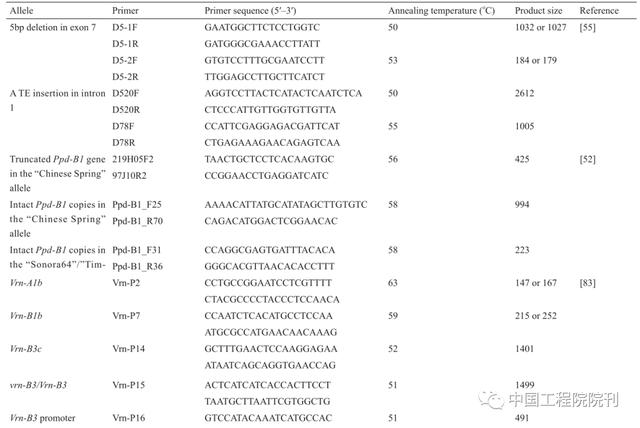
表3 不同春花和光周期反应等位基因的聚合酶链反应(PCR)标记
这些物候基因之间的复杂互作产生了两个相互矛盾的开花调控网络模型,第一个模型被命名为Vrn2-FT,在这个模型中,Vrn2抑制FT的表达,但是冬季春化作用可对Vrn1稍作上调,导致Vrn2的下调和FT表达的释放。然后,该FT与Ppd1互作,再次将Vrn1上调使之超过阈值,在长日照条件下开始开花。相反,根据缺乏Vrn1保持营养生长阶段(mvp)突变体不能上调FT的现象,Shimada等提出第二个模型,称为FT-Vrn2,Vrn1促进FT的转录,使Vrn2下调,从而启动开花。随后,Distelfeld和Dubcovsky对Vrn1分离和Vrn2缺失的mvp突变体进行详细的研究,发现了与先前提出的两种模型均相矛盾的证据,因此我们建议应该进行更多的研究来阐明小麦的开花网络,这可能发现更多在开花途径中相互作用的基因。
四、矮秆基因
将矮秆(Rht)基因引入谷类作物,包括小麦,是绿色革命的关键驱动力。从那以后,Rht-B1b和Rht-D1b(以前分别称为Rht1和Rht2),是世界各地小麦育种上最常用的Rht基因。这两个半矮秆基因一起产生矮秆表型,而单独与对应的Rht-B1a或Rht-D1a组合产生半矮秆植株。具有这些基因的植株不容易发生倒伏,并且更有效地将同化物分配到籽粒中。有些研究者认为这些品种产量潜力的提高仅限于有利的生长环境下。然而,这些特定的Rht基因对内源赤霉素不敏感,产生较矮的植株和较小的细胞。这些较小的细胞造成胚芽鞘较短、早期苗势较弱、叶面积较小、水分利用率较低、成苗率不高,特别是在水分有限的环境中。Rht-B1b和Rht-D1b等位基因对赤霉素的不敏感性是由于单核苷酸置换产生了翻译终止密码子TGA,降低了植物对赤霉素的反应能力。
世界上大多数的小麦都种植在没有灌溉的条件下,由于对季节性降雨的依赖,其产量常常受到水资源短缺的阻碍。大约50%的雨水可以通过土壤蒸发直接流失,而早期苗势强可以使水分利用率提高25%,从而提高产量。同样,为了在干旱地区获得产量效益,澳大利亚南部也常常推荐晚熟品种深播技术,但当矮秆/半矮秆品种播种深度超过5 cm时,成苗就会受到影响。因此,农民播种要等到播前第一场雨,在这种情况下,澳大利亚小麦每周产量损失在140~330 kg·hm–2。胚芽鞘较长的小麦品种在播种时能够较早出苗,早期苗势较强。此外,早期的苗势强和胚芽鞘较长有助于植株免受残留除草剂的毒性效应,增强其抗杂草能力,通过遮蔽减少蒸发造成的水分流失。因此,在水资源有限的环境中培育幼苗生长势旺盛和较长胚芽鞘的品种以具备更好的适应性是小麦主要的育种目标,目前在墨西哥CIMMYT就有这样一个项目正在执行。
另一方面,一些Rht基因(如Rht7、Rht8、Rht9、Rht13和Rht14)可以降低株高,而不会影响幼苗生长势以及组织对赤霉素的反应。在胁迫条件下,植株较高的品种在茎秆中储存同化物,籽粒灌浆并不完全依赖当时的同化物。在许多有利和不利的环境中进行的一些研究表明,高度在70~100 cm之间的植株的产量比那些高于或低于这个范围的植株的产量更好。因此,采用微效Rht基因的聚合或与一个赤霉素不敏感基因的组合而得到的较低株高比较理想。不同研究报道,与其他矮秆/半矮秆品种相比,利用Rht8和(或)Rht13等位基因与Rht1和(或)Rht2可以使产量达到最高。这些Rht等位基因的连锁标记使得在大群体中同时选择两个等位基因更加容易。
五、物候与矮秆基因的生理表现
籽粒产量受到特定环境中发育阶段时间长短的强烈影响,使得作物物候学成为产量生理学的关键组成部分。在生殖阶段,特别是在开花前几周至开花后数天的时期,水分胁迫在水资源有限的环境中对作物产量的影响最为显著。Passioura强调了干旱地区用水量、水利用效率和收获指数(HI)对作物产量的重要性。在干旱环境中,一大部分可用于蒸腾作用的土壤水分从贫瘠的土壤表面蒸发,从而通过限制根系水分利用和改变冠层温度来间接影响干物质积累。在这种情况下,苗期快速生长可以通过遮阴防止蒸发。此外,开花较晚的品种不断产生分蘖,直到获得生殖发育的信号,其中许多不能产生可育穗,但通过正常的蒸腾作用影响土壤中可利用的水分。
从这一点上讲,培育干旱环境中水分利用效率高的品种还应该考虑抽穗期和有效分蘖数。此外,整个生育期的用水需求也不一样,在结实和种子发育阶段需水较多。因此,通过改变作物的发育有可能提高产量。通过物候与季节性水分的可用性调节使之与作物发育阶段同步,应该是在水资源有限的环境中(如地中海气候地区)培育小麦新品种最重要的育种目标。
HI和最终的籽粒产量在很大程度上取决于开花前后的生物量、同化物向小花的运转和生育期间的供水模式。增加同化物向发育中的小花转移可能是提高HI的一个策略,这将防止小花在开花前的败育。这可以通过增加穗发育的持续时间,减少早期阶段从而发育成大穗来实现。而且,在籽粒灌浆期大穗还与旗叶一起贡献更多的光合产物,从而提高HI。研究表明,Vrn-A1的两个春性等位基因对根和营养形态(如花的生长习性)、株高和叶长的变化有显著的影响。最近观察到开花前的生长阶段与分蘖和干物质积累之间存在显著相关性。对澳大利亚小麦品种的多年多点详细研究表明,与具有两个春性等位基因的品种相比,3个Vrn1基因座中任何一个为春性等位基因的品种开花最早。同样,所有3个Vrnl基因座的春性等位基因对提早抽穗期的影响很小,表明存在上位性或超剂量的效应。这项研究还表明,与Vrn-A1或Vrn-D1相比,Vrn-B1对缩短抽穗期的影响较弱。然而,最近有研究表明Vrn-B1对籽粒产量影响最大。
因为与Rht-B1b相比Rht-D1b的叶片气孔减少,导致抽穗前蒸腾较慢,土壤水分可在以后使用,所以携带Rht-D1b基因的半矮秆品种在开花期和籽粒灌浆期高温少雨环境下比Rht-B1b更具有优势。植物通过加速叶片干燥和死亡来减少干旱胁迫下的用水量,从而导致光合产物的减少。因此,茎秆储备成为籽粒灌浆碳水化合物的重要来源。然而,Rht-B1b和Rht-D1b基因可分别使茎秆储备减少35%和39%,因此,与现代矮秆品种相比,当同化物不足时,在胁迫环境中高秆品种往往表现较好。
六、未来策略
产量关键生理决定因素的识别和精确测定的难题是提高植物耐旱性的瓶颈,其复杂的遗传控制使得进展更加困难。因此,为了解决这个复杂的问题,在生理和分子水平上改良植物性状是至关重要的。
目前使用自动化高通量植物表型设备可极大地帮助研究人员更精确和准确地进行植株表型测定。深入了解植物生理学将解析耐旱性的遗传组成部分,而分子和基因组工具将有助于鉴定耐旱性状的候选基因和QTL。生理学和分子工具的整合将为基因功能提供新的见解。为了在干旱研究中优化产量,对生长环境和基因型-环境相互作用的详细了解是至关重要的。通过将最适合的物候基因等位基因相结合,对特定环境下基因型的微调是可行的,这样可以更好地适应现有环境。
最近的研究表明,物候基因的拷贝数变异在作物适应性中也起着至关重要的作用。Ppd-B1的拷贝数增加使开花期提早,而Vrn-A1的拷贝数增加则延长春化阶段,即延迟开花。对澳大利亚小麦品种Ppd-B1的拷贝数变异的研究发现,对于具有1~4个拷贝的5个等位基因以及Ppd-B的空拷贝的品种,其中具有等位基因较少拷贝数的植株相对于较多拷贝等位基因的植株抽穗期更晚。此外,在其他研究中发现的这些基因的单倍型变异对几个产量相关性状具有影响,因此对于不同的适应性也有影响。在过去几年间在不同国家的环境条件下有几项研究试图确定Vrn1和Ppd1基因的价。
在大多数情况下,植物材料并不能涵盖自然界中所有可用的等位基因甚至不同遗传背景的同一等位基因。因此,为了获得等位基因在育种品种中的真实效果,需要培育具有不同等位基因的适应当地条件品种的适当近等基因系(NIL)群体,当然这将需要很大的努力。
早期的成功实例是Pugsley培育的Triple Dirk,用于研究Vrn1基因的等位基因,目前英国John Innes中心正在开展培育Ppd1基因不同等位基因群体的研究。考虑到当地水资源有限的环境,这项工作肯定会促进物候基因研究,优化植物发育和生产力。因此,为了优化每个等位基因/单倍型对物候基因的影响,全面深入的研究将使植物育种者能够确定每个关键生长环境中小麦的基本遗传结构,以在胁迫条件下获得更好的产量。Servin等指出,当利用分子和生理工具鉴定和证明不同性状及其调节耐旱性基因的效率时,这几个有用的性状可以通过标记辅助基因聚合方案聚集在该基础群体中。总之,应对气候变化不利影响的成败取决于目标环境的适当特征特性(土壤属性、降水年型、干旱严重程度等),然后设计一个结合有用的物候和其他耐旱性基因的适当的作物理想株型,再辅之以良好的田间管理措施。
七、结论
由于气候变化,干旱是世界农业面临的主要威胁,预测近期情况会更加恶化。小麦是世界上种植面积最广的谷类作物,对全球粮食安全至关重要。改变发育阶段和成熟期是在不影响产量的情况下实现抗旱的最好方法之一。然而,目前对于控制小麦开花期和成熟期的基因数量了解是有限的。根据从模式植物拟南芥中获得的知识,已经报道的控制开花基因有80多个,有理由相信许多新的与小麦开花和成熟相关的基因及遗传途径尚待发现。但事实上,不同的遗传途径最终会聚合、相互作用并导致花序原基因的激活,而这些促进开花的互作网络尚未明了。由于目前一些重要的研究正在进行,小麦开花基因和通路的知识将随着时间的推移而增加,并将需要计算生物学的进步来整合和解释这些信息。此外,未来的国际合作有助于将不同研究小组和学科正在进行的研究积累整合起来,而育种学家的挑战将是使这项工作整合到新的遗传组合中。
注:本文内容呈现形式略有调整,若需可查看原文。
改编原文:
M.A.N. Nazim Ud Dowla,Ian Edwards,Graham O’Hara,Shahidul Islam,Wujun Ma.Developing Wheat for Improved Yield and Adaptation Under a Changing Climate: Optimization of a Few Key Genes[J].Engineering,2018,4(4):514-522.☟ 如,请点击文末“阅读原文”
食品安全中的主要问题
庞国芳院士谈用风险分析应对“谈农药色变”
施肥要干预:资源可持续利用促进中国农业发展
战略研究:中国耕地空间格局变化对粮食生产的影响
战略研究:粮食作物产业的可持续发展
中国工程院院刊
工程造福人类
科技开创未来
微信公众号ID :CAE-Engineering
注:论文反映的是研究成果进展,不代表《中国工程科学》杂志社的观点。
,

免责声明:本文仅代表文章作者的个人观点,与本站无关。其原创性、真实性以及文中陈述文字和内容未经本站证实,对本文以及其中全部或者部分内容文字的真实性、完整性和原创性本站不作任何保证或承诺,请读者仅作参考,并自行核实相关内容。文章投诉邮箱:anhduc.ph@yahoo.com