植物抗逆性的测定讨论与分析(原创解读植物中顺式调控序列的重要性及其鉴定与未来挑战)
前两期我们从 The Plant Cell在线发表的综述 "Cis-regulatory sequences in plants: Their importance, discovery, and future challenges" 中,学习了顺式调控元件/模块的概念和特征,以及如何鉴定并验证顺式调控模块。这期小编将接着解读顺式调控模块的在基因组中的分布及其作用机制等。

CRM 的基因组位置:基因组大小和物种的影响
根据CRM(顺式调控模块)与其邻近编码基因转录起始位点(TSS)的线性距离,可将其分成不同的类型,一般包括与基因区域(核心启动子,5’UTR,外显子、内含子和3’UTR)存在重叠的、距离TSS小于2-5 kb的、转录终止位点下游5 kb以内的、以及位于基因远端的CRMs。尽管大多数 CRM 位于其邻近蛋白质编码基因上游几 kbp 范围内,但许多 CRM 分布在其它位置,如内含子或目标基因上游或下游10 kbp 以外。对于一些物种,包括大多数植物在内,第一个内含子的平均长度显著长于其余内含子,这意味着CRMs的存在。有关拟南芥全基因组的实验和计算分析,揭示了第一内含子具有调控活性,尽管基因组的位置似乎比内含子大小更重要。位于大的内含子上的CRMs包括拟南芥中的 FLOWERING LOCUS C和AGAMOUS,以及玉米中的knotted1 。
基于距离的分类有些随意,但有助于概念化 CRM 的位置如何在物种内和物种间演变。当使用染色质开发性标记 CRM活性时 ,很明显,CRM与邻近基因的距离差异归因于基因组大小的不同。例如,在基因组非常小的拟南芥 (~135 Mbp) 和基因组稍大 (~373 Mbp) 的水稻中,分别有近 45% 和 ~25% 的 ACR 距离其目的基因1kb以内,而在玉米 (~2,400 Mbp) 中,大多数 ACR 与其邻近基因的距离超过1 kbp。CRM 位置和基因组大小之间的这种关系,在其它被子植物中也得到了证实。基因组大小和远端CRMs的比例之间的关系有诸多解释,最可能的解释即,在较大的基因组中,原本在较小的基因组中相邻的CRMs被转座子和重复序列分离。例如,在二穗短柄草中 (~355 Mbp) 中,一些CREs 可能存在于距离目的基因 500 bp 范围内的单个 ACR 中,但在玉米中,这些CREs可能因一个初始转座子的插入而分裂为两个 ACR,随后在进化过程中插入了更多的 TE(Figure 5A)。无论基因组大小如何,基因和可能的 CRM 在数量上都高度相关。此外,大的植物基因组具有更多的 ACRs,即更多的CRMs。然而,基因组大小为 ~150Mbp 到 ~5,000Mbp 的植物物种之间,ACRs数量的变化并不多于两倍。

远端 CRMs 通过长距离染色质互作发挥功能
远端 CRM,即增强子和/或沉默子,通过染色质互作影响其靶基因的表达。ChIA-PET 和 Hi-ChIP 实验确定了不同玉米和水稻组织中远端 CRM 和核心启动子之间数以万计的 H3K4me3-、H3K27ac-或 H3K27me3 介导的长距离染色体内互作。这些互作对在不同组织、不同发育时期和环境条件下是动态变化的。例如,在玉米中,CRM 和 b1 基因之间的互作频率与b1的表达水平呈正相关(Figure 5B)。有趣的是,需要注意玉米中约 40% 的远端 CRM 不与其邻近基因相互作用,即它们倾向于跳过至少一个基因并与更远的基因互作。这让人想起 DICE,一种远端 CRM,它是如何对玉米 bx1基因的表达进行调控。Mediator是一种转录共激活因子复合物,对动物的远端 CRM 和核心启动子之间的染色质互作中起着关键作用。同样,在拟南芥中,茉莉酸调节依赖于Mediator亚基MED25 的染色质互作,表明Mediator在植物染色质互作中也起着至关重要的作用。
远端 CRM 及其目的基因是如何在三维空间中找到彼此的呢?利用 3C 技术扫描全基因组发现,具有相似表观遗传状态的染色质区域,倾向于通过一种叫相分离的机制与彼此进行物理互作。相分离由蛋白质介导,包括 HP1α 及其植物同源 Agenet Domain Containing Protein 1 (ADCP1),以及 RNAPII 和Like Heterochromatin Protein 1 (LHP1)。由此,活性染色质(活性基因)、兼性异染色质(PcG 沉默基因)和经典异染色质(沉默 TE)形成独立的空间分离区域,称为拓扑相关结构域(TAD)。TAD 是基于染色质构象分析,并通过计算而被定义的基因组序列,与周围结构域中的序列相比,TAD内的序列彼此之间具有更高的互作频率。同时,位于同一个TAD中的DNA序列之间的互作频率高于与其它TAD中序列的互作频率。在动物中,CRM-基因互作似乎主要局限于 TAD,不同 TAD 之间的边界充当绝缘体。然而,在植物中,类似的现象并不存在,即尚未发现真正意义上的绝缘体元件,但却发现了跨域TADs的长距离染色质互作。
基于 3C 的实验已鉴定到了大量的染色质互作。例如,启动子-启动子 (P-P) 频繁发生互作,参与此类互作的基因往往倾向于共表达。P-P 互作可能归因于基因共调控,但也可能是由于具有相似染色质状态的 DNA 序列聚集而不影响彼此的表达导致的相分离引起。部分P-P 互作也可能是部分核心启动子表现出增强子活性以调控远端基因。定义远端 CRM-基因对,需要区分调控基因表达的互作与由于空间物理接近而产生的染色质互作。
转座子元件:CRMs的主要来源
TF驱动的CRMs
TE 可以通过几种不同的机制影响基因表达,包括破坏 CRM、将沉默染色质扩散到侧翼基因以及提供新的 CRM。在这里,我们关注后者。人们越来越认识到 TE 是 CRM 的来源。据估计,人类和玉米中分别约有 25% 和 30% 的调控序列源自 TE(TE-CRM)。已报道哺乳动物和植物中的几个 TE 可与 TF 结合。哺乳动物和玉米细胞中的瞬时报告基因分析表明,大量测试的 TE-CRM 确实具有增强子或沉默子活性。在玉米中,测试的十个 TE-ACRs 中有八个驱动报告基因表达。因此,尽管大多数 TE 在大多数组织中似乎是沉默的,但很大一部分可能具有调节功能。
TE-CRM 的特殊之处在于它们具有在基因组内移动和扩增的潜力,物种特异的 CRMs 通常富含 TEs。具有有利影响的新的插入在进化上是保守的,其可在进化过程中重新建立基因调控网络和稳定新的基因网络,而具有不利影响的插入可能会被突变中和。例如,人类胚胎干细胞和拟南芥中分别与哺乳动物妊娠和种子发育相关的基因表达调控网络,已经被证实是重新建立的。一般,按比例来说,"年龄"较大的 TE 似乎比"年轻"的 TE 更频繁地显示调控活性。这个概念即,TE 越老,CRM 失去 DNA 甲基化并为宿主行使调控功能的时间就越长。值得注意的是,某些 TE 家族,例如内源性人类和小鼠逆转录病毒,表现出比其他家族更大的调控潜力。在植物中,在绝对数量上,更多的 CRM 位于逆转录病毒中而不是 DNA 转座子中。然而,在分析一系列植物物种中这两类转座元件的富集情况时,CRM 在 DNA 转座子中的富集程度高于预期,尤其是 hAT 亚类。值得注意的是,TE-CRM 的数量可能被严重低估。这是由于大多数全基因组分析仅保留比对上参考基因组的 unique reads,这使得reads难以比对到重复的 TE 序列。此外,与其他 CRM 一样,仅在分析相关组织、细胞类型和条件时,才可检测到 TE 充当组织、细胞类型或应激特异性的 CRM。
TE相关CRMs的实例
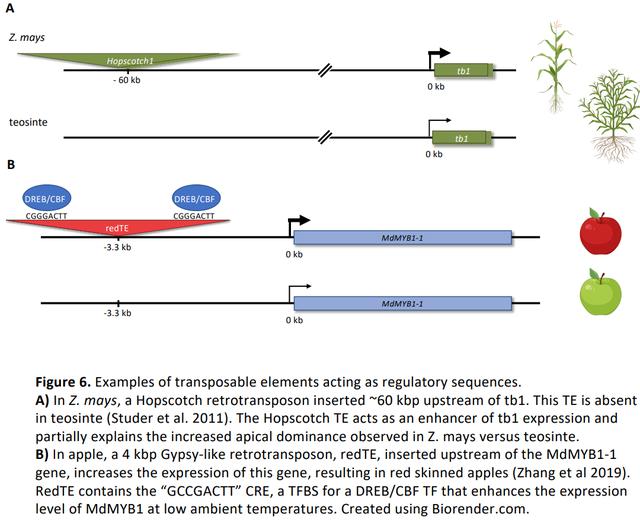
尽管全基因组分析表明多个植物基因组中存在丰富的 TE-CRM,然而,与TE-CRMs相关的某种特异表型的典型例子相对较少。一个经典的例子是与野生近缘种 teosinte 相比,Hopscotch逆转录转座子提高了其靶基因 tb1 在驯化玉米中的表达水平(Figure 6A)。有研究通过 STARR-seq 从部分Hopscotch元件中检测到了增强子活性。在拟南芥中,在 Jacalin Lectin 家族蛋白基因的启动子中删除四分之一的 TE,可以将这些基因从根特异性表达转变为组成型表达模式,表明删除的 TE 具有沉默子活性。 TE 破坏 CRM 功能的一个例子是 Vgt1中MITETE 的插入,Vgt1是玉米花抑制基因ZmRap2.7的增强子。这种 MITE 插入与早花性状有关。
对于许多 TE-CRMs,参与调控相关靶基因的 TFBS 是已知。例如,在苹果中,一个 4 kb 的反转录转座子 redTE 增加了MdMYB1 的表达水平,导致果皮颜色变红(Figure 6B)。RedTE包含dehydration-responsive element/C-repeat-binding (DREB/CBF) TF 的结合motif,可在相对较低的环境温度下增强MdMYB1 的表达。有趣的是,在玉米中,与冷、热和盐诱导表达基因相关的TE 也被证实富含 DREB/CBF TFBS。类似地,拟南芥中的 ONSEN 转座子具有热响应 CREs,可招募热激转录因子 A2 (HSFA2) 以激活邻近基因。
尽管 DNA 甲基化模式在植物组织中非常稳定,但有研究表明,在各种胁迫条件下,TE-CRM 的 DNA 去甲基化可能会增强特异的胁迫响应基因的表达。例如,RESISTANCE METHYLATED GENE 1的表达与位于DNA 去甲基化酶 REPRESSOR OF SILENCING 1 (ROS1)编码区域上游的两个Helitron相关 TE 重复序列的去甲基化有关。这些结果表明 TE-CRMs 的 DNA 去甲基化可能在植物免疫反应中发挥作用。目前尚不清楚观察到的 TE 侧翼序列的 DNA 甲基化变化是否与连锁的应激反应基因的上调存在因果关系。不过,现已证实,基因表达的变化会导致附近 TE 的 DNA 甲基化水平发生变化。
植物中CRMs序列的保守性
CRM 的序列多样化是物种间和物种内表型变异的关键因素。CRM 的功能多样化,以及基因和基因调控网络的功能多样化,是由物种杂交和全基因组复制驱动的。CRMs 的其中一个重复可能丢失,或者两个重复都多样化,导致了新功能化和亚功能化。的确,研究重复CRMs随时间变化的命运表明,CRMs在旁系同源物之间的保留与重复事件的发生时间呈负相关。
随着时间的推移,研究重复 CRM 的命运表明,旁系同源物之间 CRM 的保留与重复事件的发散时间呈负相关。 对玉米和大豆中旁系同源远端 ACRs 的分析表明,其中大约 50%(在大豆中 >50%),都可以在所研究的组织中获得。在物种对中,例如玉米-高粱和大豆-菜豆,超过一半的远端 CRMs 在这些物种对中共有,其中三分之二在两个物种中染色质都是开放的(Figure 7)。重复 CRMs 的多样化受各种因素影响,包括驯化、选择、基因组重复的时间以及序列丢失和变化的速度。
CRMs 序列多样化的速度取决于它们的功能。许多 CRMs 在进化过程中是保守的,表明它们具有重要的功能;比较分析可用于识别此类 CRMs。一致认为,中枢神经系统富含染色质开放区域和增强子。使用比较方法识别 CRMs 早期依赖于位点特定的系统发育树,由此近缘物种被用来寻找进化产生的遗传差异,以识别重要的保守区域。这种方法在植物中鉴定了出多种 CRMs,例如拟南芥中的 AG 和FLOWERING LOCUS T(FT) 基因座,以及芸苔属中的MIR164基因座。AG 基因的内含子甚至在几种十字花科植物中包含多个保守的 TF motif。系统发育树也在全基因组中得以应用,可通过比较同线直系同源序列来发现保守的非编码序列 (CNS)。同线性通常是识别全基因组 CNS 的重要手段。
尽管 CNS 可用于识别保守和多样化的 CRMs,但核苷酸替换率可能会限制 CRM 的检测。因此,许多 CRM 将需要其他数据来确定其位置,例如染色质结构数据。总之,关于 CRMs 序列保守性的信息,支持这一观点,即非编码序列变异导致表型变异,伴随着进化过程中植物多样化和适应性的产生。

未来的挑战和展望
现在是研究植物基因组顺式调控序列的俱佳时期,然而仍有不少困难需要克服。其一是对植物组织和条件特异性CRMs的鉴定,单细胞测序技术可以克服之一困难。单细胞RNA-seq和ATAC-seq技术已成功应用于植物研究,然而单细胞ChIP和Hi-C和WGBS技术在动物中得以突破,目前在植物中的应用尚未见报道。尽管如此,毋庸置疑的是,不同类型的单细胞测序数据是揭示植物基因组顺式调控组的利器。其二是如何鉴定CRMs是增强子而不是沉默子或者两者皆是,填补沉默子的知识空白是解决这一问题的有效方法。其三是如何鉴定CRM-基因对,由于大量CRMs并非调控其邻近基因的表达,使其变得更为困难。基于单细胞3C的技术极具挑战,幸运的是,单细胞ATAC-seq结合单细胞RNA-seq可以揭示共同开放的ACRs和靶基因。最近一项研究揭示玉米叶片细胞类型中约78%的Hi-C互作对应于CRM-基因对。另外,鉴定TFs及其共作用因子也面临较大的挑战。
目前研究顺式调控元件正慢慢变得白热化,但未来我们的注意力应该转向这些知识的应用,可以结合基因编辑技术,往提高植物的适应性方向努力。
原文链接
https://academic.oup.com/plcell/advance-article/doi/10.1093/plcell/koab281/6433166?searchresult=1
关注我们
小麦族多组学网站:http://202.194.139.32
投稿、合作等邮箱:13148474750@163.com
微信群:
论文查重、润色
杂志影响因子查询:
,
免责声明:本文仅代表文章作者的个人观点,与本站无关。其原创性、真实性以及文中陈述文字和内容未经本站证实,对本文以及其中全部或者部分内容文字的真实性、完整性和原创性本站不作任何保证或承诺,请读者仅作参考,并自行核实相关内容。文章投诉邮箱:anhduc.ph@yahoo.com






